Почему модель хищник жертва является не точной. Курсовая работа: Качественное исследование модели хищник-жертва. по дисциплине «Моделирование систем»
Федеральное агентство по образованию
Государственное образовательное учреждение
высшего профессионального образования
«Ижевский государственный технический университет»
Факультет «Прикладная математика»
Кафедра «Математическое моделирование процессов и технологий»
по дисциплине «Дифференциальные уравнения»
Тема: «Качественное исследование модели хищник-жертва»
Ижевск 2010
ВВЕДЕНИЕ
1. ПАРАМЕТРЫ И ОСНОВНОЕ УРАВНЕНИЕ МОДЕЛИ «ХИЩНИК-ЖЕРТВА»
2.2 Обобщенные модели Вольтера типа «хищник-жертва».
3. ПРАКТИЧЕСКОЕ ПРИМЕНЕНИЕ МОДЕЛИ «ХИЩНИК-ЖЕРТВА»
ЗАКЛЮЧЕНИЕ
СПИСОК ЛИТЕРАТУРЫ
ВВЕДЕНИЕ
В настоящее время задачи экологии имеют первостепенное значение. Важным этапом решения этих задач является разработка математических моделей экологических систем.
Одной из основных задач экологии па современном этапе является изучение структуры и функционирования природных систем, поиск общих закономерностей. Большое влияние на экологию оказала математика, способствующая становлению математической экологии, особенно такие её разделы, как теория дифференциальных уравнений, теория устойчивости и теория оптимального управления.
Одной из первых работ в области математической экологии была работа А.Д. Лотки (1880 - 1949), который первый описал взаимодействие различных популяций, связанных отношениями хищник - жертва. Большой вклад в исследование модели хищник -жертва внесли В. Вольтерра (1860 - 1940), В.А. Костицин (1883-1963) В настоящее время уравнения описывающие взаимодействие популяций, называются уравнениями Лотки - Вольтерра.
Уравнения Лотки - Вольтерра описывают динамику средних величин - численности популяции. В настоящее время на их основе построены более общие модели взаимодействия популяций, описываемые интегро-дифференциальными уравнениями, исследуются управляемые модели хищник - жертва.
Одной из важных проблем математической экологии является проблема устойчивости экосистем, управления этими системами. Управление может осуществляться с целью перевода системы из одного устойчивого состояния в другое, с целью её использования или восстановления.
1. ПАРАМЕТРЫ И ОСНОВНОЕ УРАВНЕНИЕ МОДЕЛИ ХИЩНИК-ЖЕРТВА
Попытки математического моделирования динамики как отдельных биологических популяций, так и сообществ, включающих взаимодействующие популяции различных видов, предпринимались давно. Одна из первых моделей роста изолированной популяции (2.1) была предложена еще в 1798 г. Томасом Мальтусом:
Данная модель задается следующими параметрами:
N - численность популяции;
Разность между коэффициентами рождаемости и смертности.
Интегрируя это уравнение получаем:
![]() , (1.2)
, (1.2)
где N(0) – численность популяции в момент t = 0. Очевидно, что модель Мальтуса при > 0 дает бесконечный рост численности, что никогда не наблюдается в природных популяциях, где ресурсы, обеспечивающие этот рост, всегда ограничены. Изменения численности популяций растительного и животного мира нельзя описывать простым законом Мальтуса, на динамику роста влияют многие взаимосвязанные причины – в частности, размножение каждого вида саморегулируется и видоизменяется так, чтобы этот вид сохранялся в процессе эволюции.
Математическим описанием этих закономерностей занимается математическая экология – наука об отношениях растительных и животных организмов и образуемых ими сообществ между собой и с окружающей средой.
Наиболее серьезное исследование моделей биологических сообществ, включающих в себя несколько популяций различных видов, было проведено итальянским математиком Вито Вольтерра:
 ,
,
где - численность популяции;
Коэффициенты естественного прироста (или смертности) популяции; - коэффициенты межвидового взаимодействия. В зависимости от выбора коэффициентов модель описывает либо борьбу видов за общий ресурс, либо взаимодействие типа хищник - жертва, когда один вид является пищей для другого. Если в работах других авторов основное внимание уделялось построению различных моделей, то В. Вольтерра провел глубокое исследование построенных моделей биологических сообществ. Именно с книги В. Вольтерра, по мнению многих ученых, началась современная математическая экология.
2. КАЧЕСТВЕННОЕ ИССЛЕДОВАНИЕ ЭЛЕМЕНТАРНОЙ МОДЕЛИ «ХИЩНИК-ЖЕРТВА»
2.1 Модель трофического взаимодействия по типу «хищник-жертва»
Рассмотрим модель трофического взаимодействия по типу «хищник-жертва», построенную В. Вольтерром. Пусть имеется система, состоящая из двух видов, из которых один поедает другой.
Рассмотрим случай, когда один из видов является хищником, а другой - жертвой, и будем считать, что хищник питается только жертвой. Примем следующую простую гипотезу:
Коэффициент прироста жертвы;
Коэффициент прироста хищника;
Численность популяции жертвы;
Численность популяции хищника;
Коэффициент естественного прироста жертвы;
Скорость потребления жертвы хищником;
Коэффициент смертности хищника в отсутствие жертвы;
Коэффициент «переработки» хищником биомассы жертвы в собственную биомассу.
Тогда динамика численности популяций в системе хищник - жертва будет описываться системой дифференциальных уравнений (2.1):
 (2.1)
(2.1)
где все коэффициенты положительные и постоянные.
Модель имеет равновесное решение (2.2):
По модели (2.1) доля хищников в общей массе животных выражается формулой (2.3):
 (2.3)
(2.3)
Анализ устойчивости состояния равновесия по отношению к малым возмущениям показал, что особая точка (2.2) является «нейтрально» устойчивой (типа «центр»), т. е. любые отклонения от равновесия не затухают, но переводят систему в колебательный режим с амплитудой, зависящей от величины возмущения. Траектории системы на фазовой плоскости имеют вид замкнутых кривых, расположенных на различных расстояниях от точки равновесия (рис. 1).

Рис. 1 – Фазовый «портрет» классической вольтерровой системы «хищник-жертва»
Разделив первое уравнение системы (2.1) на второе, получим дифференциальное уравнение (2.4) для кривой на фазовой плоскости .
 (2.4)
(2.4)
Интегрируя данное уравнение получим:
![]() (2.5)
(2.5)
где - постоянная интегрирования, где 
Несложно показать, что движение точки по фазовой плоскости будет происходить только в одну сторону. Для этого удобно сделать замену функций и , перенеся начало координат на плоскости в стационарную точку (2.2) и введя затем полярные координаты:
 (2.6)
(2.6)
В таком случае, подставив значения системы (2.6) в систему (2.1), будем иметь:
 (2.7)
(2.7)
Умножив первое уравнение на , а второе - на и сложив их, получим:
После аналогичных алгебраических преобразований получим уравнение для :
Величина , как видно из (4.9), всегда больше нуля. Таким образом, не меняет знака, и вращение все время идет в одну сторону.
Интегрируя (2.9) найдем период:
Когда мало, то уравнения (2.8) и (2.9) переходят в уравнения эллипса. Период обращения в этом случае равен:
 (2.11)
(2.11)
Исходя из периодичности решений уравнений (2.1), можно получить некоторые следствия. Представим для этого (2.1) в виде:
 (2.12)
(2.12)
и проинтегрируем по периоду:
 (2.13)
(2.13)
Так как подстановки от и в силу периодичности обращаются в нуль, средние по периоду оказываются равными стационарным состояниям (2.14):
 (2.14)
(2.14)
Простейшие уравнения модели «хищник-жертва» (2.1) обладают рядом существенных недостатков. Так, в них предполагается неограниченность пищевых ресурсов для жертвы и неограниченный рост хищника, что противоречит экспериментальным данным. Кроме того, как видно из рис. 1, ни одна из фазовых кривых не выделена с точки зрения устойчивости. При наличии даже небольших возмущающих воздействий траектория системы будет все дальше уходить от положения равновесия, амплитуда колебаний расти, и система достаточно быстро разрушится.
Несмотря на недостатки модели (2.1), представления о принципиально колебательном характере динамики системы «хищник- жертва» получили широкое распространение в экологии. Взаимодействиями «хищник-жертва» объясняли такие явления, как колебания численности хищных и мирных животных в промысловых зонах, колебания в популяциях рыб, насекомых и т. д. На самом деле колебания численности могут быть обусловлены и другими причинами.
Предположим, что в системе хищник - жертва происходит искусственное уничтожение особей обоих видов, и рассмотрим вопрос о том, каким образом уничтожение особей влияет на средние значения их численности, если осуществляется пропорционально этой численности с коэффициентами пропорциональности и соответственно для жертвы и хищника. С учетом сделанных предположений систему уравнений (2.1) перепишем в виде:
 (2.15)
(2.15)
Предположим, что , т. е. коэффициент истребления жертвы меньше коэффициента ее естественного прироста. В этом случае также будут наблюдаться периодические колебания численности. Вычислим средние значения численностей:
 (2.16)
(2.16)
Таким образом, если , то средняя численность популяций жертвы возрастает, а хищника - убывает.
Рассмотрим случай, когда коэффициент истребления жертвы больше коэффициента ее естественного прироста, т. Е . В этом случае ![]() при любых , и, следовательно, решение первого уравнения (2.15) ограничено сверху экспоненциально убывающей функцией
при любых , и, следовательно, решение первого уравнения (2.15) ограничено сверху экспоненциально убывающей функцией ![]() , т. е. при .
, т. е. при .
Начиная с некоторого момента времени t, при котором , решение второго уравнения (2.15) также начинает убывать и при стремится к нулю. Таким образом, в случае оба вида исчезают.
2.1 Обобщенные модели Вольтера типа «хищник-жертва»
Первые модели В. Вольтерра, естественно, не могли отражать все стороны взаимодействия в системе хищник - жертва, поскольку они были в значительной мере упрощены относительно реальных условий. Например, если численность хищника равна нулю, то из уравнений (1.4) следует, что численность жертвы неограниченно возрастает, что не соответствует действительности. Однако ценность этих моделей состоит именно в том, что они были основой, на которой быстрыми темпами начала развиваться математическая экология.
Появилось большое число исследований различных модификаций системы хищник - жертва, где были построены более общие модели, учитывающие в той или иной степени реальную ситуацию в природе.
В 1936 г. А.Н. Колмогоров предложил использовать для описания динамики системы хищник - жертва следующую систему уравнении:
 , (2.17)
, (2.17)
где убывает с возрастанием численности хищников, а возрастает с увеличением численности жертвы.
Эта система дифференциальных уравнений в силу ее достаточной общности позволяет хорошо учитывать реальное поведение популяций и вместе с тем проводить качественный анализ ее решений.
Позднее в своей работе, Колмогоров исследовал подробно менее общую модель:
 (2.18)
(2.18)
Различные частные случаи системы дифференциальных уравнений (2.18) исследовались многими авторами. В таблице приведены различные частные случаи функций , , .
Таблица 1 - Различные модели сообщества «хищник-жертва»
| Авторы | |||
| Вольтерра-Лотка | |||
| Гаузе | |||
| Пислоу | |||
| Холинг | |||
| Ивлев | |||
| Рояма | |||
| Шимазу | |||
| Мэй |
математическое моделирование хищник жертва
3. ПРАКТИЧЕСКОЕ ПРИМЕНЕНИЕ МОДЕЛИ ХИЩНИК-ЖЕРТВА
Рассмотрим математическую модель совместного существования двух биологических видов (популяций) типа "хищник - жертва", называемую моделью Вольтерра - Лотки.
Пусть два биологических вида совместно обитают в изолированной среде. Среда стационарна и обеспечивает в неограниченном количестве всем необходимым для жизни один из видов, который будем называть жертвой. Другой вид - хищник также находится в стационарных условиях, но питается лишь особями первого вида. Это могут быть караси и щуки, зайцы и волки, мыши и лисы, микробы и антитела и т. д. Будем для определенности называть их карасями и щуками.
Заданы следующие начальные показатели:
Со временем число карасей и щук меняется, но так как рыбы в пруду много, то не будем различать 1020 карасей или 1021 и поэтому будем считать и непрерывными функциями времени t. Будем называть пару чисел (,) состоянием модели.
Очевидно, что характер изменения состояния (,) определяется значениями параметров. Изменяя параметры и решая систему уравнений модели, можно исследовать закономерности изменения состояния экологической системы во времени.
В экосистеме скорость изменения численности каждого вида также будем считать пропорциональной его численности, но только с коэффициентом, который зависит от численности особей другого вида. Так, для карасей этот коэффициент уменьшается с увеличением числа щук, а для щук увеличивается с увеличением числа карасей. Будем считать эту зависимость также линейной. Тогда получим систему из двух дифференциальных уравнений:

Эта система уравнений и называется моделью Вольтерра-Лотки. Числовые коэффициенты , , - называются параметрами модели. Очевидно, что характер изменения состояния (,) определяется значениями параметров. Изменяя эти параметры и решая систему уравнений модели, можно исследовать закономерности изменения состояния экологической системы.
Проинтегрируем оба уравнения систему по t, которое будет изменяться от - начального момента времени, до , где T – период, за который происходят изменения в экосистеме. Пусть в нашем случае период равен 1 году. Тогда система принимает следующий вид:
 ;
;
 ;
;
Принимая = и = приведем подобные слагаемые, получим систему, состоящую из двух уравнений:

Подставив в полученную систему исходные данные получим популяцию щук и карасей в озере спустя год:
Хищники могут поедать травоядных животных, и также слабых хищников. Хищники обладают широким спектром питания, легко переключаются с одной добычи на другую, более доступную. Хищники часто нападают на слабые жертвы. Поддерживается экологическое равновесие между популяциями жертва-хищник.[ ...]
Если равновесие неустойчиво (предельных циклов нет) или внешний цикл неустойчив, то численности обоих видов, испытывая сильные колебания, уходят из окрестности равновесия. Причем быстрое вырождение (в первой ситуации) наступает при низкой адаптации хищника, т.е. при его высокой смертности (по сравнению со скоростью размножения жертвы). Это означает, что слабый во всех отношениях хищник не способствует стабилизации системы и сам вымирает.[ ...]
Пресс хищников особенно силен, когда в коэволюции хищник - жертва равновесие смещается в сторону хищника и ареал жертвы сужается. Конкурентная борьба тесно связана с нехваткой пищевых ресурсов, она может быть и прямой борьбой, например, хищников за пространство как ресурс, но чаще всего это просто вытеснение вида, которому на данной территории пищи не хватает, видом, которому этого же количества пищи вполне достаточно. Это уже межвидовая конкуренция.[ ...]
 |
И окончательно в системе ’’хищник - жертва”, описываемой моделью (2.7), возникновение диффузионной неустойчивости (при локальной устойчивости равновесия) возможно лишь в том случае, когда естественная смертность хищника возрастает с ростом его численности быстрее, чем линейная функция, и трофическая функция отличается от вольтерровской либо, когда популяция жертвы - это популяция типа Олли.[ ...]
Теоретически в моделях «один хищник - две жертвы» эквивалентное выедание (отсутствие предпочтения того или иного вида жертвы) может повлиять на конкурентное сосуществование видов-жертв лишь в тех местах, где уже существует потенциально устойчивое равновесие . Разнообразие может возрасти только в таких условиях, когда у видов с меньшей конкурентоспособностью скорость роста популяции выше, чем у доминантов. Это позволяет понять ситуацию, когда равномерное выедание ведет к увеличению видового разнообразия растений там, где большее число видов, прошедших отбор на быстрое размножение, сосуществует с видами, эволюция которых направлена на повышение конкурентоспособности .[ ...]
Точно так же выбор жертвы, зависящий от ее плотности, может привести к устойчивому равновесию в теоретических моделях двух конкурирующих видов жертвы, где прежде никакого равновесия не существовало . Для этого хищник должен был бы обладать способностью к функциональным и численным реакциям на изменения плотности жертвы; возможно, однако, что переключение (непропорционально частые нападения на наиболее обильную жертву) будет иметь при этом более важное значение. В самом деле, установлено, что переключение оказывает стабилизирующее влияние в системах «один хищник - п жертв» и представляет собой единственный механизм, способный стабилизировать взаимодействия в тех случаях, когда ниши жертв полностью перекрываются . Такую роль могут играть неспециализированные хищники. Предпочтение более специализированными хищниками доминантного конкурента действует таким же образом, как переключение хищника, и может стабилизировать теоретические взаимодействия в моделях, в которых прежде не существовало равновесия между видами жертвы, при условии что их ниши в какой-то степени разделены .[ ...]
Также- не стабилизирует сообщество и хищник ’’сильный во всех отношениях”, т.е. хорошо адаптированный к данной жертве и с низкой относительной смертностью. При этом система обладает неустойчивым предельным циклом и, несмотря на устойчивость положения равновесия, вырождается в случайной среде (хищник выедает жертву и вследствие этого гибнет). Такая ситуация соответствует медленному вырождению.[ ...]
Таким образом, при хорошей адаптации хищника в окрестности устойчивого равновесия могут возникать неустойчивый и устойчивый циклы, т.е. в зависимости от начальных условий система ’’хищник-жертва” либо стремится к равновесию, либо, колеблясь, уходит от него, либо в окрестности равновесия устанавливаются устойчивые колебания численностей обоих видов.[ ...]
Организмы, которых относят к хищникам, питаются другими организмами, уничтожая свою жертву. Таким образом, среди живых организмов следует выделять еще одну классификационную систему, а именно «хищники» и «жертвы». Отношения между такими организмами складывались на протяжении всей эволюции жизни на нашей планете. Организмы-хищники выступают в роли природных регуляторов численности организмов-жертв. Увеличение численности «хищников» приводит к уменьшению численности «жертв», это, в свою очередь, снижает запасы пищи («жертв») для «хищников», что в целом диктует снижение численности «жертв» и т. д. Таким образом, в биоценозе постоянно происходят колебания численности хищников и жертв, в целом же устанавливается определенное равновесие на какой-то период времени в пределах достаточно устойчивых условий среды.[ ...]
Это в конечном итоге приходит к экологическому равновесию между популяциями хищника и жертвы.[ ...]
Для трофической функции третьего типа состояние равновесия будет устойчивым, если где N - точка перегиба функции (см. рис. 2, в). Это следует из того, что на интервале трофическая функция вогнута и, следовательно, относительная доля потребления хищником жертвы возрастает.[ ...]
Пусть Гг = -Г, т.е. имеет место сообщества типа ’’хищник - жертва”. В этом случае первое слагаемое в выражении (7.4) равно нулю и для выполнения условия устойчивости по вероятности равновесного состояния N требуется, чтобы и второе слагаемое не было положительным.[ ...]
Таким образом, для рассмотренного сообщества типа хищник - жертва можно сделать вывод об асимптотической устойчивости в целом положительного положения равновесия, т. е. для любых начальных данных 1Ч(0)>0 эволюция происходит таким образом, что N(7) - ■ К при условии, что N >0.[ ...]
Так, в однородной среде, не имеющей укрытий для размножения, хищник рано или поздно уничтожает популяцию жертвы и после этого вымирает сам. Волны жизни” (изменений численности хищника и жертвы) следуют друг за другом с постоянным сдвигом по фазе, и в среднем численность как хищника, так и жертвы остается примерно на одном уровне. Длительность периода зависит от скоростей роста обоих видов и от исходных параметров. Для популяции жертвы влияние хищника положительно, так как ее чрезмерное размножение привело бы к краху ее численности. В свою очередь, все механизмы, препятствующие полному истреблению жертвы, способствуют сохранению пищевой базы хищника.[ ...]
Другие модификации могут быть следствием особенностей поведения хищника. Число особей жертвы, которое хищник в состоянии потребить в данное время, имеет свой предел. Эффект насыщения хищника при приближении к этому рубежу показан в табл. 2-4, В. Взаимодействия, описываемые уравнениями 5 и 6, могут иметь устойчивые точки равновесия или обнаруживать циклические колебания. Однако такие циклы отличаются от тех, какие отражены в уравнениях Лотки - Вольтерры 1 и 2. Циклы, передаваемые уравнениями 5 и 6, могут иметь постоянную амплитуду и средние плотности, пока среда постоянна; после того как произошло нарушение, они могут вернуться к прежним амплитудам и средним плотностям. Такие циклы, которые восстанавливаются после нарушений, называются устойчивыми предельными циклами. Взаимодействие зайца и рыси можно считать устойчивым предельным циклом, но это не цикл Лотки - Вольтерры.[ ...]
Рассмотрим возникновение диффузионной неустойчивости в системе ’’хищник -жертва”, но сначапа выпишем условия, обеспечивающие возникновение диффузионной неустойчивости в системе (1.1) при п = 2. Ясно, что равновесие (N , Щ) локально (т.е.[ ...]
Перейдем к интерпретации случаев, связанных с длительным сосуществованием хищника и жертвы. Понятно, что в отсутствие предельных циклов устойчивому равновесию будут соответствовать в случайной среде флуктуации численности, причем их амплитуда будет пропорциональна дисперсии возмущений. Такое явление будет происходить, если хищник имеет высокую относительную смертность и в то же время высокую степень приспособленности к данной жертве.[ ...]
Рассмотрим теперь, как меняется динамика системы с ростом приспособленности хищника, т.е. с убыванием Ъ от 1 до 0. Если приспособленность достаточно низкая, то предельные циклы отсутствуют, а равновесие является неустойчивым. С ростом приспособленности в окрестности этого равновесия возможно появление устойчивого цикла и далее внешнего неустойчивого. В зависимости от начальных условий (соотношения биомассы хищника и жертвы) система может либо терять устойчивость, т.е. уходить из окрестности равновесия, либо в ней будут со временем устанавливаться устойчивые колебания. Дальнейший рост приспособленности делает невозможным колебательный характер поведения системы. Однако при Ъ [ ...]
Пример отрицательной (стабилизирующей) обратной связи - взаимоотношение между хищником и жертвой или функционирование карбонатной системы океана (раствор СОг в воде: СО2 + Н2О -> Н2СОз). Обычно количество углекислоты, растворенное в воде океана, находится в парциальном равновесии с концентрацией углекислого газа в атмосфере. Локальные увеличения углекислоты в атмосфере после извержения вулканов приводят к интенсификации фотосинтеза и поглощению ее карбонатной системой океана. При снижении уровня углекислого газа в атмосфере карбонатная система океана высвобождает СОг в атмосферу. Поэтому концентрация углекислого газа в атмосфере достаточно стабильна.[ ...]
[ ...]
Как отмечает Р.Риклефс (1979), существуют факторы, способствующие стабилизации взаимоотношений в системе “хищник-жертва”: неэффективность хищника, наличие у хищника альтернативных пищевых ресурсов, уменьшение запаздывания в реакции хищника, а также экологические ограничения, налагаемые внешней средой на ту или иную популяцию. Взаимодействия между популяциями хищника и жертвы весьма разнообразны и сложны. Так, если хищники достаточно эффективны, они могут регулировать плотность популяции жертвы, удерживая ее на уровне ниже емкости среды. Через влияние, оказываемое ими на популяции жертвы, хищники воздействуют на эволюцию различных признаков жертвы, что приводит в конечном итоге к экологическому равновесию между популяциями хищника и жертвы.[ ...]
Если выполняется одно из условий: 0 1/2. Если же 6 > 1 (кА [ ...]
Устойчивость биоты и окружающей среды зависит только от взаимодействия растений - автотрофов и растительноядных гетеротрофных организмов. Хищники любых размеров не способны нарушить экологическое равновесие сообщества, так как в естественных условиях они не могут увеличить свою численность при постоянной численности жертв. Хищники не только должны быть сами передвигающимися, но и могут питаться только передвигающимися животными.[ ...]
Никакие другие рыбы не распространены так широко, как щуки. В немногих местах товли в стоячих или проточных водоемах нет давления со стороны щук для сохранения равновесия между жертвой и хищником Только современные искусственные водоемы, в которых щуки являются рыбами нежелательными из-за разведения там других рыб, не заселяются ими целенаправленно. В мире щуки исключительно хорошо представлены. Их ловят по всему северном) полушарию от Соединенных Штатов и Канады в Северной Америке, через Европу до северной части Азии.[ ...]
Еще одна возможность устойчивого сосуществования возникает здесь же, в узком диапазоне сравнительно высокой адаптации. При переходе к неустойчивому режиму с очень ’’хорошим” хищником может возникнуть устойчивый внешний предельный цикл, при котором диссипация биомассы уравновешивается ее притоком в систему (высокой продуктивностью жертвы). Тогда возникает любопытная ситуация, когда наиболее вероятными являются два характерных значения амплитуды случайных колебаний. Одни происходят вблизи равновесия, другие - вблизи предельного цикла, причем возможны более или менее частые переходы между этими режимами.[ ...]
Гипотетические популяции, которые ведут себя в соответствии с векторами на рис. 10.11 А, изображены на рис. 10.11,-Б с помощью графика, показывающего динамику соотношения численностей хищника и жертвы и на рис. 10.11,5 в виде графика динамики численности хищника и жертвы во времени. В популяции жертвы, по мере того как она переходит от равновесия прн низкой плотности к равновесию при высокой и возвращается назад, происходит «вспышка» численности. И эта вспышка численности не является следствием столь же выраженного изменения в окружающей среде. Напротив, это изменение численности порождено самим воздействием (при небольшом уровне «шума» в среде) и оно, в частности, отражает существование нескольких равновесных состояний. Сходные рассуждения можно использовать для объяснения более сложных случаев динамики численности в природных популяциях.[ ...]
Важнейшим свойством экосистемы является ее устойчивость, сбалансированность обмена и происходящих в ней процессов. Способность популяций или экосистемы поддерживать устойчивое динамическое равновесие в изменяющихся условиях среды называется гомеостазом (homoios - тот же, подобный; stasis - состояние). В основе гомеостаза лежит принцип обратной связи. Для поддержания равновесия в природе не требуется внешнего управления. Пример гомеостаза - субсистема «хищник-жертва», в которой регулируются плотность популяций хищника и жертвы.[ ...]
Природная экосистема (биогеоценоз) устойчиво функционирует при постоянном взаимодействии ее элементов, круговороте веществ, передаче химической, энергетической, генетической и другой энергии и информации по цепям-каналам. Согласно принципу равновесия любая естественная система с проходящим через нее потоком энергии и информации имеет тенденцию к развитию устойчивого состояния. При этом устойчивость экосистем обеспечивается автоматически за счет механизма обратной связи. Обратная связь заключается в использовании получаемых данных от управляемых компонентов экосистем для внесения корректив управляющими компонентами в процесс. Рассмотренные выше взаимоотношения «хищник» - «жертва» в данном контексте можно описать несколько подробнее; так, в водной экосистеме хищные рыбы (щука в пруду) поедают другие виды рыб-жертвы (карась); если численность карася будет увеличиваться -это пример положительной обратной связи; щука, питаясь карасем, снижает его численность-это пример отрицательной обратной связи; при росте числа хищников снижается число жертв, и хищник, испытывая недостаток пищи, также снижает рост своей популяции; в конце концов в рассматриваемом пруду устанавливается динамическое равновесие в численности и щуки, и карася. Постоянно поддерживается равновесие, которое исключало бы исчезновение любого звена трофической цепи (рис. 64).[ ...]
Переходим к наиболее важному обобщению, а именно что отрицательные взаимодействия со временем становятся менее заметными, если экосистема достаточно стабильна и ее пространственная структура обеспечивает возможность взаимного приспособления популяций. В модельных системах типа хищник- жертва, описываемых уравнением Лотки-Вольтерры, если в уравнение не введены дополнительные члены, характеризующие действие факторов самоограничения численности, то колебания происходят непрерывно и не затухают (см. Левонтин, 1969). Пиментел (1968; см. также Пиментел и Стоун, 1968) экспериментально показал, что такие дополнительные члены могут отражать взаимные адаптации или генетическую обратную связь. Когда же новые культуры создавали из особей, ранее на протяжении двух лет совместно существовавших в культуре, где их численность подвергалась значительным колебаниям, оказалось, что у них выработался экологический гомеостаз, при котором каждая из популяций была «подавлена» другой в такой степени, что оказалось возможным их сосуществование при более стабильном равновесии.
КОМПЬЮТЕРНАЯ МОДЕЛЬ «ХИЩНИК-ЖЕРТВА»
Казачков Игорь Алексеевич 1 , Гусева Елена Николаевна 2
1 Магнитогорский государственный технический университет им. Г.И. Носова, институт строительства, архитектуры и искусства, студент 5 курса
2 Магнитогорский государственный технический университет им. Г.И. Носова, институт энергетики и автоматизированный систем, кандидат педагогических наук, доцент кафедры бизнес-информатики и информационных технологий
Аннотация
Данная статья посвящена обзору компьютерной модели «хищник-жертва». Проведенное исследование позволяет утверждать, что экологическое моделирование играет огромную роль в исследовании окружающей среды. Данная проблематика имеет многогранный характер.
COMPUTER MODEL «PREDATOR-VICTIM»
Kazatchkov Igor Alekseevich 1 , Guseva Elena Nikolaevna 2
1 Nosov Magnitogorsk State Technical University, Civil Engineering, Architecture and Arts Institute, student of the 5th course
2 Nosov Magnitogorsk State Technical University, Power Engineering and Automated Systems Institute, PhD in Pedagogical Science, Associate Professor of the Business Computer Science and Information Technologies Department
Abstract
This article provides an overview of the computer model "predator-victim". The study suggests that environmental simulation plays a huge role in the study of the environment. This problem is multifaceted.
Для исследования окружающей нас среды используют экологическое моделирование. Математические модели используют в тех случаях, когда нет естественной среды и нет естественных объектов, она помогает сделать прогноз влияния разных факторов на исследуемый объект. Данный метод берет на себя функции проверки, построения и интерпретацию полученных результатов. На основе таких форм экологическое моделирование занимается оценкой изменений, окружающей нас среды.
В настоящий момент подобные формы используется для изучения окружающей нас среды, а когда требуется изучить какую-либо из ее областей, то применяют математическое моделирование. Данная модель дает возможность спрогнозировать влияние тех или иных факторов на объект изучения. В свое время был предложен тип «хищник – жертва» такими учеными как: Т. Мальтусом (Malthus 1798, Мальтус 1905), Ферхюльстом (Verhulst 1838), Пирлом (Pearl 1927, 1930), а также А. Лотки (Lotka 1925, 1927) и В. Вольтерры (Volterra 1926).Эти модели воспроизводят периодический колебательный режим, возникающий в результате межвидовых взаимодействий в природе.
Одним из основных методов познания является моделировка. Помимо того, что в нем можно спрогнозировать изменения, происходящие в окружающей среде, к тому же помогает найти оптимальный способ решения проблемы. Уже давно в экологии используют математические модели, для того чтобы установить закономерности, тенденции развития популяций, помогают выделить суть наблюдений. Макет может служить образцом поведения, объекта.
При воссоздании объектов в математической биологии используются прогнозирования различных систем, предусматриваются специальные индивидуальности биосистем: внутренне строение особи, условия жизнеобеспечения, постоянство экологических систем, благодаря которым сберегается жизнедеятельность систем.
Появление компьютерного моделирования значительно раздвинуло рубеж способностей исследования. Возникло вероятность многосторонней реализации трудных форм, не допускающих аналитического изучения, появились новейшие направления, а еще имитационное моделирование.
Рассмотрим, что же такое объект моделирования. «Объектом является замкнутая среда обитания, где происходит взаимодействие двух биологических популяций: хищников и жертв. Процесс роста, вымирания и размножения происходит
непосредственно на поверхности среды обитания. Питание жертв происходит за счет тех ресурсов, которые присутствуют в данной среде, а питание хищников происходит за счет жертв. При этом питательные ресурсы могут быть как возобновляемые, так и не возобновляемые.
В 1931 году Вито Вольтеррой были выведены следующие законы отношения хищник-жертва.
Закон периодического цикла – процесс уничтожения жертвы хищником нередко приводит к периодическим колебаниям численности популяций обоих видов, зависящим только от скорости роста плотоядных и растительноядных, и от исходного соотношения их численности.
Закон сохранения средних величин – средняя численность каждого вида постоянна, независимо от начального уровня, при условии, что специфические скорости увеличения численности популяций, а также эффективность хищничества постоянны.
Закон нарушения средних величин – при сокращении обоих видов пропорционально их числу, средняя численность популяции жертвы растет, а хищников – падает.
Модель хищник-жертва – это особая взаимосвязь хищника с жертвой, в результате которой выигрывают оба. Выживают наиболее здоровые и приспособленные особи к условиям среды обитания, т.е. все это происходит благодаря естественному отбору. В той среде где нет возможности для размножения, хищник рано или поздно уничтожит популяцию жертвы, в последствии чего вымрет и сам» .
На земле существует множество живых организмов, которые при благоприятных условиях увеличивают численность сородичей до огромных масштабов. Такая способность называется: биотический потенциал вида, т.е. увеличение численности вида за определенный промежуток времени. Каждый вид имеет свой биотический потенциал, к примеру крупные виды организмов за год могут возрасти всего в 1,1 раза, в свою очередь организмы более мелких видов, таких как рачки и т.д. могут увеличить свой вид до 1030 раз, ну а бактерии еще в большем количестве. В любом из этих случаев популяция будет расти в геометрической прогрессии.
Экспоненциальным ростом численности называется геометрическая прогрессия роста численности популяции. Такую способность можно наблюдать в лаборатории у бактерий, дрожжей. В не лабораторных условиях экспоненциальный рост возможно увидеть на примере саранчи или же на примере других видов насекомых. Такой рост численности вида можно наблюдать в тех местах где у него практически нет врагов, а продуктов питания более чем достаточно. В конце концов увеличение вида, после того как численность возросла в течении непродолжительного времени, рост популяции начинал снижаться.
Рассмотрим компьютерную модель размножения млекопитающих на примере модели Лотки-Вольтерры. Пусть на некоторой территории обитают два вида животных: олени и волки. Математическая модель изменения численности популяций в модели
Лотки-Вольтерры:
Начальное число жертв - xn, число хищников - yn.
Параметры модели:
P1– вероятность встречи с хищником,
P2– коэффициент роста хищников за счет жертв,
d – коэффициент смертности хищников,
a – коэффициент прироста численности жертв.
В учебной задаче были заданы такие значения: численность оленей равнялось 500, численности волков равна 10, коэффициент прироста оленей равен 0,02, коэффициент прироста численности волков равен 0,1, вероятность встречи с хищником 0,0026, коэффициент роста хищников за счет жертв 0,000056. Данные рассчитаны на 203 года.
Исследуем влияние коэффициент прироста жертв на развитие двух популяций, остальные параметры оставим без изменений.
На схеме 1 наблюдается увеличение численности жертвы и затем, с некоторым опозданием наблюдается прирост хищников. Затем хищники выбивают жертв, число жертв резко падает и вслед за ним уменьшается число хищников (рис. 1).

Рисунок 1. Численность популяций при низкой рождаемости у жертв
Проанализируем изменение модели, увеличив коэффициент рождаемости жертвы а=0,06. На схеме 2 мы видим циклический колебательный процесс, приводящий к увеличению численности обоих популяций со временем (рис. 2).
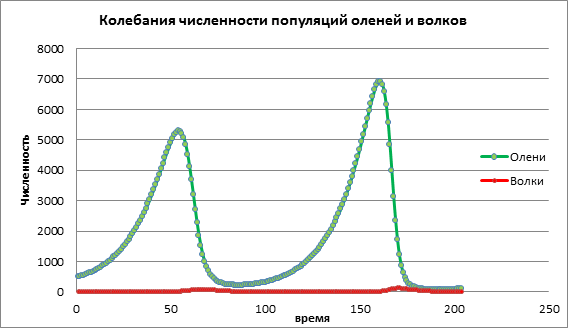
Рисунок 2.Численность популяций при средней рождаемости у жертв
Рассмотрим как изменится динамика популяций при высоком значении коэффициента рождаемости жертвы а=1,13. На рис. 3 наблюдается резкое увеличение численности обеих популяций с последующим вымиранием, как жертвы, так и хищника. Это происходит за счет того, что численность популяции жертв увеличилось до такого количества, что стали заканчиваться ресурсы, вследствие чего происходит вымирание жертвы. Вымирание хищников происходит из-за того, что сократилось количество жертв и у хищников закончились ресурсы для существования.
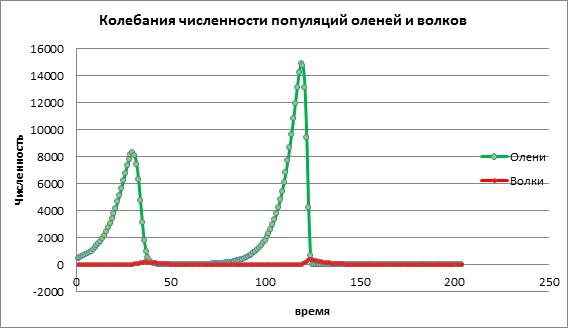
Рисунок 3.Численность популяций при высокой рождаемости у жертв
Исходя из анализа данных компьютерного эксперимента, можно сделать выводы о том, что компьютерное моделирование позволяет нам прогнозировать численность популяций, изучать влияние различных факторов на популяционную динамику. В приведенном примере мы исследовали модель «хищник-жертва», влияние коэффициента рождаемости жертв на численность оленей и волков. Небольшой прирост популяции жертв приводит к небольшому увеличению жертв, которую через некоторый период уничтожают хищники. Умеренный прирост популяции жертв приводит к увеличению численности обеих популяций. Высокий прирост популяции жертв приводит сначала к быстрому росту популяции жертв, это влияет на увеличение роста хищников, но затем расплодившиеся хищники быстро уничтожают популяцию оленей. В итоге оба вида вымирают.
В модели Колмогорова сделано одно существенное допущение: поскольку предполагается, что при то это означает существование в популяции жертв механизмов, регулирующих их численность даже в отсутствие хищников.
К сожалению, подобная формулировка модели не позволяет ответить на вопрос, вокруг которого в последнее время ведется много споров и о котором мы уже упоминали в начале главы: каким образом популяция хищников может оказывать регулирующее воздействие на популяцию жертв с тем, чтобы вся система была устойчива? Поэтому мы вернемся к модели (2.1), в которой механизмы саморегуляции (например, регулирование при помощи внутривидовой конкуренции) в популяции жертв (впрочем, как и в популяции хищников) отсутствуют; поэтому единственным механизмом регуляции численностей, входящих в сообщество видов, являются трофические взаимоотношения между хищниками и жертвами.
Здесь (так что, в отличие от предыдущей модели, Естественно, что решения (2.1) зависят от конкретного вида трофической функции которая, в свою очередь, определяется характером хищничества, т. е. трофической стратегией хищника и защитной стратегией жертвы. Общими для всех этих функций (см. рис. И) являются следующие свойства:
Система (2.1) имеет одну нетривиальную стационарную точку, координаты которой определяются из уравнений
![]()
при естественном ограничении .
Существует еще одна стационарная точка (0, 0), соответствующая тривиальному равновесию. Нетрудно показать, что эта точка - седло, а сепаратрисами являются координатные оси.
Характеристическое уравнение для точки имеет вид
![]()
Очевидно, что для классической вольтерровской модели .
Поэтому величину f можно рассматривать как меру отклонения рассматриваемой модели от вольтерровской.
![]()
стационарная точка - фокус, и в системе появляются колебания; при выполнении противоположного неравенства - узел, и колебания в системе отсутствуют. Устойчивость этого состояния равновесия определяется условием
т. е. существенным образом зависит от вида трофической функции хищника.
Условие (5.5) можно интерпретировать следующим образом: для устойчивости нетривиального равновесия системы хищник - жертва (и тем самым, для существования этой системы) достаточно, чтобы в окрестности этого состояния относительная доля потребляемых хищником жертв возрастала с возрастанием численности жертв. Действительно, доля жертв (от общей их численности), потребляемых хищником, описывается дифференцируемой функцией условие возрастания которой (положительность производной) выглядит как
![]()
Последнее условие, взятое в точке есть не что иное, как условие (5.5) устойчивости равновесия. При непрерывности оно должно выполняться и в некоторой окрестности точки Таким образом, если численности жертв в этой окрестности, то
Пусть теперь трофическая функция V имеет вид, изображенный на рис. 11, а (характерный для беспозвоночных). Можно показать, что для всех конечных значений (поскольку выпукла вверх)
т. е. при любых величинах стационарной численности жертв неравенство (5.5) не выполняется.
Это означает, что в системе с таким видом трофической функции не существует устойчивого нетривиального равновесия. Возможны несколько исходов: либо численности как жертвы, так и хищника неограниченно возрастают, либо (при прохождении траектории вблизи одной из координатных осей) в силу случайных причин численность жертвы или численность хищника станет равной нулю. При гибели жертвы через некоторое время погибнет и хищник, если же сначала погибает хищник, то численность жертвы начнет экспоненциально возрастать. Третий вариант - возникновение устойчивого предельного цикла - невозможен, что легко доказывается.
В самом деле, выражение

в положительном квадранте всегда положительно, если только имеет вид, изображенный на рис. 11, а. Тогда, согласно критерию Дюлака в этой области отсутствуют замкнутые траектории и устойчивого предельного цикла существовать не может.
Итак, можно сделать вывод: если трофическая функция имеет вид, изображенный на рис. 11, а, то хищник не может быть регулятором, обеспечивающим стабильность популяции жертвы и тем самым устойчивость всей системы в целом. Система может быть устойчива только в том случае, когда в популяции жертвы существуют собственные внутренние регулирующие механизмы, например, внутривидовая конкуренция или эпизоотии. Этот вариант регулирования был уже рассмотрен в §§ 3, 4.
Ранее отмечалось, что такой тип трофической функции характерен для хищников-насекомых, «жертвами» которых также обычно являются насекомые. С другой стороны, наблюдения за динамикой многих природных сообществ типа «хищник - жертва», включающих в себя виды насекомых, показывают, что для них характерны колебания очень большой амплитуды и весьма специфического вида.
Обычно после более или менее постепенного роста численности (который может происходить либо монотонно, либо в виде колебаний с возрастающей амплитудой) происходит ее резкое падение (рис. 14), и затем картина повторяется. По-видимому, такой характер динамики численностей видов насекомых может быть объяснен неустойчивостью этой системы при малых и средних значениях численности и действием мощных внутрипопуляционных регуляторов численности при больших ее значениях.

Рис. 14. Динамика численности популяции австралийской листоблошки Cardiaspina albitextura, кормящейся на эвкалиптах. (Из статьи: Clark L. R. The population dynamics of Cardiaspina albitextura.-Austr. J. Zool., 1964, 12, № 3, p. 362-380.)
Если же в систему «хищник - жертва» входят виды, способные к достаточно сложному поведению (например, хищники способны к обучению или жертвы способны находить убежище), то в такой системе возможно существование устойчивого нетривиального равновесия. Доказывается это утверждение весьма просто.
В самом деле, трофическая функция тогда должна иметь вид, изображенной на рис. 11, в. Точка на этом графике - точка касания прямой, проведенной из начала координат, графика трофической функции Очевидно, что в этой точке функция имеет максимум. Также легко показывается, что для всех выполняется условие (5.5). Следовательно, нетривиальное равновесие, в котором численность жертв меньше будет асимптотически устойчиво
Однако мы ничего не можем сказать о том, сколь велика область устойчивости этого равновесия. Например, если существует неустойчивый предельный цикл, то эта область должна лежать внутри цикла. Или другой вариант: нетривиальное равновесие (5.2) неустойчиво, но существует устойчивый предельный цикл; в этом случае также можно говорить о стабильности системы хищник - жертва. Поскольку выражение (5.7) при выборе трофической функции типа рис. 11, в может менять знак при изменении при , то критерий Дюлака здесь не работает и вопрос о существовании предельных циклов остается открытым.

Хищничество - форма трофических взаимоотношений между организмами разных видов, при которых один из них (хищник ) атакует другого (жертву ) и питается его плотью, то есть обычно присутствует акт умерщвления жертвы.
Система «хищник - жертва» - сложная экосистема, для которой реализованы долговременные отношения между видами хищника и жертвы, типичный пример коэволюции.
Коэволюция - совместная эволюция биологических видов, взаимодействующих в экосистеме.
Отношения между хищниками и их жертвами развиваются циклически, являясь иллюстрацией нейтрального равновесия.
1. Единственным лимитирующим фактором, ограничивающим размножение жертв, является давление на них со стороны хищников. Ограниченность ресурсов среды для жертвы не учитывается.
2. Размножение хищников ограничивается количеством добытой им пищи (количеством жертв).
По своей сути модель Лотки–Вольтерры есть математическое описание дарвинского принципа борьбы за существование.
Система Вольтерра-Лотка, которую часто называют системой хищники-жертвы, описывает взаимодействие двух популяций - хищников (например, лисиц) и жертв (например, зайцев), которые живут по несколько разным "законам". Жертвы поддерживают свою популяцию за счет поедания природного ресурса, например, травы, что приводит к экспоненциальному росту численности популяции, если нет хищников. Хищники поддерживают свою популяцию за счет только "поедания" жертв. Поэтому, если популяция жертв исчезает, то вслед за этим популяция хищников экспоненциально убывает. Поедание хищниками жертв наносит ущерб популяции жертв, но в то же время дает дополнительный ресурс к размножению хищников.
Вопрос
ПРИНЦИП МИНИМАЛЬНОГО РАЗМЕРА ПОПУЛЯЦИЙ
закономерно существующее в природе явление, характеризуемое как своеобразный естественный принцип,означающий, что каждому виду животных свойствен специфический для него минимальный размерпопуляции, нарушение которого ставит под угрозу существование популяции, а иногда и вида в целом.
правило популяционного максимума, оно заключается в том, что популяция не может увеличиваться бесконечно, в силу истощения пищевых ресурсов и условий размножения (теория андреварты – бирча) и ограничения воздействия комплекса абиотических и биотических факторов среды (теория фредерикса).
Вопрос
Итак, как было ясно уже Фибоначчи, прирост популяции пропорционален ее численности, и поэтому, если рост популяции не ограничивают никакие внешние факторы, он непрерывно ускоряется. Опишем этот рост математически.
Прирост популяции пропорционален численности особей в ней, то есть ΔN~N , где N - численность популяции, а ΔN - ее изменение за определенный период времени. Если этот период бесконечно мал, можно написать, что dN/dt=r ×N , где dN/dt - изменение численности популяции (прирост), а r - репродуктивный потенциал , переменная, характеризующая способность популяции увеличивать свою численность. Приведенное уравнение называется экспоненциальной моделью роста численности популяции (рис. 4.4.1).
Рис.4.4.1. Экспоненциальный рост .
Как легко понять, с ростом времени численность популяции растет все быстрее, и достаточно скоро устремляется к бесконечности. Естественно, никакое местообитание не выдержит существования популяции с бесконечной численностью. Тем не менее, существует целый ряд процессов популяционного роста, который в определенном временном промежутке может быть описан с помощью экспоненциальной модели. Речь идет о случаях нелимитированного роста, когда какая-то популяция заселяет среду с избытком свободного ресурса: коровы и лошади заселяют пампу, мучные хрущаки - элеватор с зерном, дрожжи - бутыль виноградного сока и т.д.
Естественно, экспоненциальный рост популяции не может быть вечным. Рано или поздно ресурс исчерпается, и рост популяции затормозится. Каким будет это торможение? Практическая экология знает самые разные варианты: и резкий взлет численности с последующим вымиранием популяции, исчерпавшей свои ресурсы, и постепенное торможение прироста по мере приближения к определенному уровню. Проще всего описать медленное торможение. Простейшая описывающая такую динамику модель называется логистической и предложена (для описания роста численности популяции человека) французским математиком Ферхюльстом еще в 1845 году. В 1925 году аналогичная закономерность была заново открыта американским экологом Р. Перлем, который предположил, что она носит всеобщий характер.
В логистической модели вводится переменнаяK - емкость среды , равновесная численность популяции, при которой она потребляет все имеющиеся ресурсы. Прирост в логистической модели описывается уравнением dN/dt=r ×N ×(K-N)/K (рис. 4.4.2).

Рис. 4.4.2. Логистический рост
Пока N невелико, на прирост популяции основное влияние оказывает сомножитель r ×N и рост популяции ускоряется. Когда становится достаточно высоким, на численность популяции начинает оказывать основное влияние сомножитель (K-N)/K и рост популяции начинает замедляться. Когда N=K , (K-N)/K=0 и рост численности популяции прекращается.
При всей своей простоте логистическое уравнение удовлетворительно описывает много наблюдаемых в природе случаев и до сих пор с успехом используется в математической экологии.
№16.Экологическая стратегия выживания - эволюционно сложившийсякомплекс свойств популяции, направленный на повышение вероятности выживания и оставление потомства.
Так А.Г. Раменский (1938) различал три основные типа стратегий выживания среди растений: виоленты, патиенты и эксплеренты.
Виоленты (силовики) - подавляют всех конкурентов, например, деревья, образующие коренные леса.
Патиенты - виды, способные выжить в неблагоприятных условиях («тенелюбивые», «солелюбивые» и т.п.).
Эксплеренты (наполняющие) - виды, способные быстро появляться там, где нарушены коренные сообщества, - на вырубках и гарях (осины), на отмелях и т.д.
Экологические стратегии популяций отличаются большим разнообразием. Но при этом все их многообразие заключено между двумя типами эволюционного отбора, которые обозначаются константами логистического уравнения: r-стратегия и К-стратегия.
| Признак | r-стратегии | K-стратегии |
| Смертность | Не зависит от плотности | Зависит от плотности |
| Конкуренция | Слабая | Острая |
| Продолжительность жизни | Короткая | Долгая |
| Скорость развития | Быстрое | Медленное |
| Сроки размножения | Ранние | Поздние |
| Репродуктивное усиление | Слабое | Большое |
| Тип кривой выживания | Вогнутая | Выпуклая |
| Размер тела | Мелкий | Крупный |
| Характер потомства | Много, мелкие | Мало, крупные |
| Размер популяций | Сильные колебания | Постоянный |
| Предпочитаемая среда | Изменчивая | Постоянная |
| Стадии сукцессии | Ранние | Поздние |
Похожая информация.

